
Sciences de la Vie et de la Terre 1 Bac
La communication nerveuse
Cours (Partie 2)
Professeur : Mr BAHSINA Najib
Sommaire
III- Nature et naissance du message nerveux
3-1/ Mise en évidence du potentiel de repos
3-2/ Mise en évidence du potentiel d’action
3-3/ Les phénomènes ioniques à l’origine du potentiel de repos
3-4/ Les phénomènes ioniques à l’origine du potentiel d’action
III- Nature et naissance du message nerveux
3-1/ Mise en évidence du potentiel de repos
Pour mettre en évidence l’activité électrique du nerf au repos, on utilise des techniques d’expérimentations récentes, un support vivant favorable tel que des fibres nerveuses géantes de Calmar, dont le diamètre de l’axone atteint 1mm et une cuve contenant un liquide nutritif isotonique :
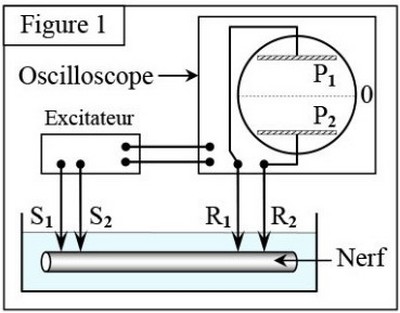
III- Nature et naissance du message nerveux
3-1/ Mise en évidence du potentiel de repos
En absence de toute excitation, on réalise les deux expériences suivantes :
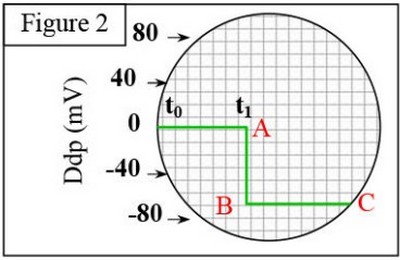
1- Au début, on place les deux microélectrodes et à la surface de l’axone, on observe sur l’écran la ligne 0A
2- Au temps , on introduit, progressivement, la microélectrode à l’intérieur de l’axone ( reste à la surface), on obtient la partie ABC, sur le même enregistrement .
III- Nature et naissance du message nerveux
3-1/ Mise en évidence du potentiel de repos
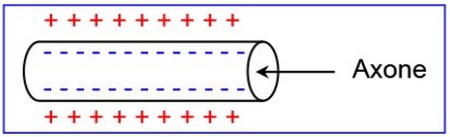
Le nerf (ou la fibre nerveuse) à l’état de repos, est caractérisé par une différence de potentiel permanente de part et d’autre de sa membrane. C’est le potentiel de repos ou potentiel membranaire.
En absence de toute stimulation, le potentiel de repos est de -7 Mv.
Il caractérise toutes les cellules vivantes
il est due à une polarité électrique entre l’intérieur et l’extérieur de ces cellules :
la surface porte des charges positives et l’intérieur des charges négatives.
III- Nature et naissance du message nerveux
3-2/ Mise en évidence du potentiel d’action
En utilisant le même montage que celui utilisé pour mesurer le potentiel de repos, on peut enregistrer l’activité électrique du nerf (ou la fibre nerveuse) à la suite d’une excitation efficace et isolée :
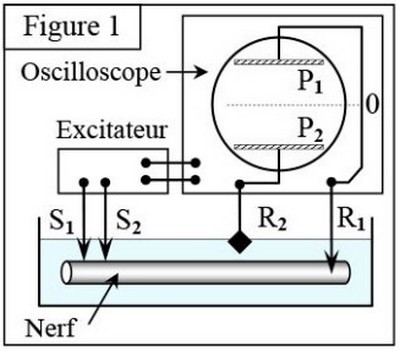
III- Nature et naissance du message nerveux
3-2/ Mise en évidence du potentiel d’action
La forme des enregistrements obtenus varie selon l’emplacement des électrodes réceptrices ( et ) :
Expérience 1
Au temps , l’électrode est introduite à l’intérieur de l’axone et une électrode de référence.
Au temps , on applique une excitation efficace.
On obtient le tracé de la figure 2 :
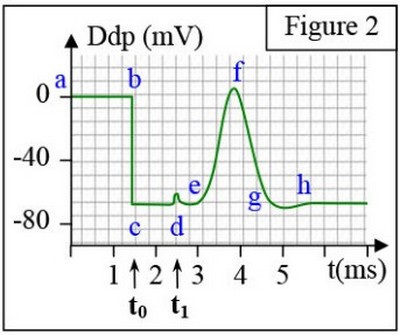
Le potentiel d’action monophasique est constitué des phases suivantes :
- L’artefact de stimulation (d).
- La phase de latence ((d - e).
- La dépolarisation au niveau de R1 (e- f).
- La repolarisation au niveau de R1(f- g).
- La phase d’hyperpolarisation (g- h).
III- Nature et naissance du message nerveux
3-2/ Mise en évidence du potentiel d’action
Expérience 2
Au temps , les électrodes et sont à la surface de l’axone.
Au temps , on applique une excitation efficace. On obtient l’enregistrement de la figure 3 :
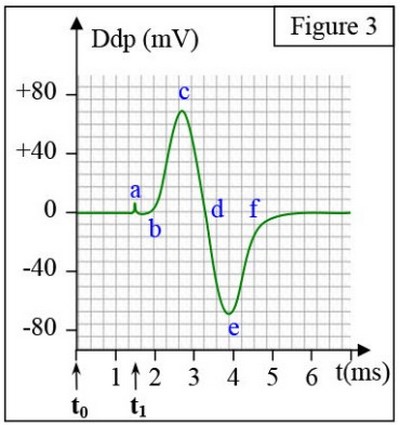
Le potentiel d’action diphasique est constitué des phases suivantes :
- L’artefact de stimulation (a).
- La phase de latence ((a - b).
- La dépolarisation au niveau de R2 (b- c).
- La repolarisation au niveau de R2 (c- d).
- La dépolarisation au niveau de R1 (d- e).
- La repolarisation au niveau de R1 (e- f).
La stimulation supraliminaire du nerf ou la fibre nerveuse en un point précis, provoque une variation transitoire du potentiel, c’est le potentiel d’action.
Selon l’emplacement des électrodes réceptrices ( et ), le potentiel d’action peut être soit :
- Un potentiel d’action monophasique .
- Un potentiel d’action diphasique .
III- Nature et naissance du message nerveux
3-2/ Mise en évidence du potentiel d’action
L’excitation efficace du nerf ou la fibre nerveuse en un point précis, provoque une inversion temporaire de la différence de potentiel, de part et d’autre de la membrane cellulaire, appelée potentiel d’action, qui se propage sous forme d’une onde de négativité formant le message nerveux :
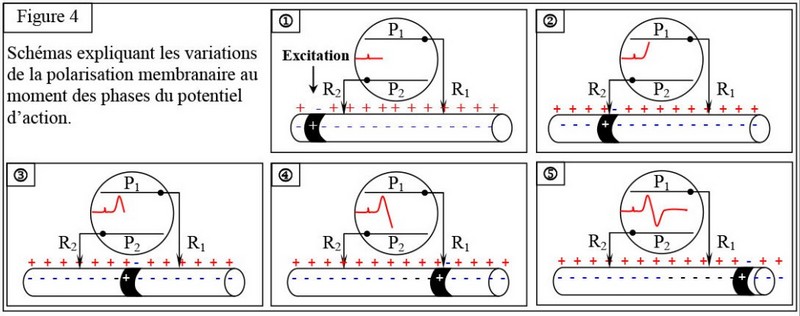
Lorsque cette onde arrive à l’électrode réceptrice , elle crée une dépolarisation de cette électrode.
Lorsque cette onde quitte l’électrode réceptrice, il y’aura rétablissement du potentiel membranaire de repos.
Cette étape est appelée phase de repolarisation de l’électrode réceptrice.
III- Nature et naissance du message nerveux
3-3/ Les phénomènes ioniques à l’origine du potentiel de repos
Origine du potentiel de repos
Pour connaître l’origine du potentiel de repos, on réalise les expériences suivantes :
Expérience 1 :
On mesure la répartition des ions Na" et K+ de part et d’autre de la membrane cytoplasmique de l’axone.
Les résultats de cette étude sont présentés par le tableau suivant :
| Ions | Intérieur de l’axone en mg/L | Extérieur de l’axone en mg/L |
| 50 | 440 | |
| 410 | 22 |
La répartition des ions de part et d’autre de la membrane de la fibre nerveuse est inégale.
Le milieu externe est riche en ion et pauvre en ions ,
le milieu intérieur est riche en ions et pauvre en ions .
L’existence d’un gradient de concentration d’une espèce ionique de part et d’autre de la membrane entraîne l’existence d’un gradient électrique.
Le potentiel de repos peut-être expliqué par la répartition dissymétrique des ions de part et d’autre de la membrane, donc de la perméabilité sélective de la membrane aux ions.
III- Nature et naissance du message nerveux
3-3/ Les phénomènes ioniques à l’origine du potentiel de repos
Origine du potentiel de repos
Expérience 2 :
Un axone amyélinique géant de calmar est plongé dans le liquide de Ringer (Maintient en vie un organe animal isolé) contenant le sodium radioactif .
Après quelques heures dans ce milieu, on replonge cet axone dans du liquide contenant des ions Na+ non radioactives.
Les résultats de cette expérience sont présentés par la figure suivante :
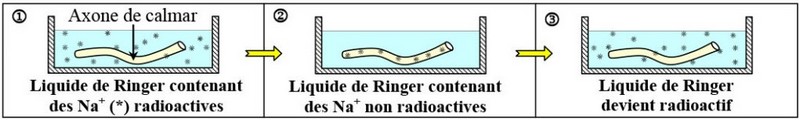
1- Phase 1 : La solution extérieure contient le radioactif. Passage du radioactif du milieu extérieur vers le milieu intérieur de Faxone.
2- Phase 2 : Le milieu extérieur contient le normal. L’axone contient le radioactif.
3- Phase 3 : Passage de radioactif de l’intérieur de l’axone vers le milieu extérieur.
Les ions tendent à diffuser selon leur gradient de concentration.
Si les échanges des ions se font uniquement par transport passif suivant le gradient de concentration, le potentiel de repos tend à disparaître progressivement.
Donc il existe un autre mécanisme qui agit pour conserver la différence de concentration des ions de part et d’autre de la membrane de la fibre nerveuse.
III- Nature et naissance du message nerveux
3-3/ Les phénomènes ioniques à l’origine du potentiel de repos
Transport actif et maintien du potentiel de repos
On mesure la sortie de sodium radioactif d’un axone géant de calmar en fonction du temps, en conditions normales, puis en ajoutant un inhibiteur de la synthèse d'ATP : le dinitrophénol (DNP).
On utilise le sodium radioactif, pour pouvoir le détecter et donc savoir ce qu'il devient une fois introduit dans l'axone.
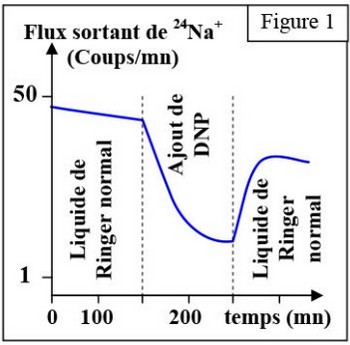
La figure 1, présente les résultats de cette expérience :
III- Nature et naissance du message nerveux
3-3/ Les phénomènes ioniques à l’origine du potentiel de repos
Transport actif et maintien du potentiel de repos
Au début de l'expérience, on s'aperçoit que le flux sortant de sodium radioactif diminue régulièrement avec le temps.
Cela montre que le introduit dans l'axone est progressivement évacué de la cellule.
Après l’ajout de DNP, on constate une nette diminution de ce flux qui n'est que temporaire puisqu'une fois le DNP retiré du milieu, le flux sortant de sodium retrouve sa pente initiale.
C'est donc qu'en conditions normales le sodium sort de la cellule et que cette sortie nécessite de l'ATP.
III- Nature et naissance du message nerveux
3-3/ Les phénomènes ioniques à l’origine du potentiel de repos
Transport actif et maintien du potentiel de repos
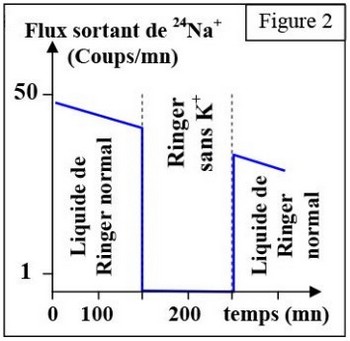
On mesure la sortie de sodium radioactif d’un axone géant de calmar en fonction du temps, en conditions normales, puis en ajoutant les ions .
La figure 2, présente les résultats de cette expérience :
III- Nature et naissance du message nerveux
3-3/ Les phénomènes ioniques à l’origine du potentiel de repos
Transport actif et maintien du potentiel de repos
On constate que le flux sortant de sodium radioactif s’arrête brusquement, une fois le retiré du milieu.
Cela montre que la sortie des ions nécessite la présence des ions .
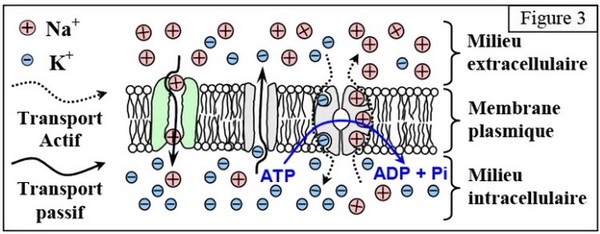
La sortie de sodium nécessitant de l'ATP et la présence des ions , il s'agit donc d'un mécanisme de transport actif et on peut penser qu'il s'agit de la pompe :
III- Nature et naissance du message nerveux
3-3/ Les phénomènes ioniques à l’origine du potentiel de repos
Conclusion
Le potentiel de repos résulte d’une inégale répartition des ions et entre le milieu extérieur, du nerf et le milieu intérieur.
Cette différence de concentration s’explique par des échanges ioniques qui se font à travers la membrane cellulaire sous l’effet de deux types de transport :
- Un transport passif, qui se fait selon le gradient décroissant de la concentration.
- Un transport actif, qui se fait contre le gradient de concentration, par des protéines membranaires spécifiques connues sous le nom de pompe à , qui nécessite de l’énergie.
Au moment de chaque cycle fonctionnel de la membrane plasmique, la pompe à fait sortir 3 ions pour entrer 2 ions , avec l’hydrolyse d’une molécule d’ATP selon la réaction :
III- Nature et naissance du message nerveux
3-4/ Les phénomènes ioniques à l’origine du potentiel d’action
Origine du potentiel d’action
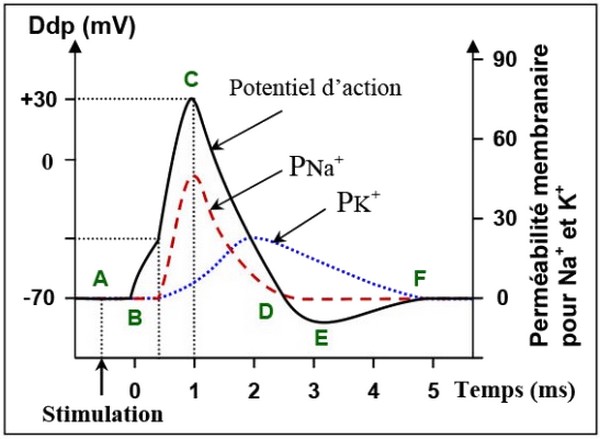
Pour déterminer l’origine du potentiel d’action, on porte une stimulation efficace sur l’axone et on mesure en fonction du temps la variation de la perméabilité membranaire aux ions et .
Les résultats de cette expérience sont présentés sur la figure suivante :
III- Nature et naissance du message nerveux
3-4/ Les phénomènes ioniques à l’origine du potentiel d’action
Origine du potentiel d’action
- (A- B)
Au repos la perméabilité membranaire aux ions sodium et potassium est très faible car la plupart des canaux ioniques sont fermés.
Or, ces canaux étant sensibles au potentiel de membrane, une légère dépolarisation suffit à provoquer leur ouverture.
- (B- C)
L’excitation de la membrane cytoplasmique provoque instantanément et localement une augmentation de la perméabilité de la membrane cytoplasmique aux ions par ouverture locale des canaux de .
Les ions sodium rentrent alors massivement dans la cellule en raison de leur gradient de concentration et de leur gradient électrique ce qui augmente la dépolarisation et finit par inverser le potentiel de membrane qui atteint une valeur d'environ +30mV.
- (C- D)
Cette forte dépolarisation finit par inactiver les canaux au sodium mais induit l'ouverture de canaux au potassium, ce qui a pour effet d'augmenter la perméabilité aux ions potassium.
Les ions potassium, beaucoup plus nombreux à l'intérieur qu'à l'extérieur, quittent alors la cellule en masse et permettent au potentiel de membrane de retrouver sa valeur initiale.
- (D- E- F)
Toutefois les canaux au potassium n'étant pas immédiatement inactivés au moment où la fibre retrouve son potentiel de repos, les ions potassium continuent à quitter la cellule et provoquent ainsi une légère hyper-polarisation, le temps que la perméabilité au potassium retrouve sa valeur de repos.
Dans le même temps, la pompe s'active, en consommant de l’énergie, fait sortir , et fait entrer contre leurs gradient de concentration.
Ce sont en effet des mouvements de sodium et de potassium qui sont à l'origine des différentes phases du potentiel d'action.
III- Nature et naissance du message nerveux
3-4/ Les phénomènes ioniques à l’origine du potentiel d’action
Les structures membranaires responsables des échanges ioniques
Les structures membranaires responsables de la variation de répartition des ions à travers la membrane cellulaire au moment du potentiel d’action sont des protéines membranaires spécifiques responsables de la variation de la perméabilité membranaire vis à vie des ions et qui fonctionnent sous l’action de la différence de potentiel électrique membranaire locale provoquée par l’excitation.
On les appelle canaux voltage-dépendants.

